Most people probably wouldn’t consider bustling towns and cities good places for nature to thrive. Yet a few species of birds have so successfully adapted to city living that they boast large and thriving urban populations. Now, research has suggested that the success of these city-dwelling species may lie in their behaviour.
Urban habitats are quite different to the natural environments in which birds evolved. Cities are noisy places, they are lit almost continually with artificial lights and they contain an abundance of food. Cities are also full of people. This means birds living there rarely get any peace and must cope with almost constant disturbance from both humans and their pets.
But birds living in cities are known to be much more tolerant of human disturbance than their rural compatriots. In a study of 44 European bird species, all but four allowed humans to approach them more closely in cities than in rural habitats. This suggests that city birds are bolder in the face of a potential threat.
It’s not only towards humans that urban birds seem to be bolder. We recently studied the territorial behaviour of male great tits (Parus major) in cities and rural habitats in the UK and found significant differences between them.
During the early spring, males of this species defend breeding territories. Once a male has a good territory he signals his ownership to other birds using song. In fact, together with attracting females, one of the main uses of song is territory defence.
When a rival male enters another’s territory and begins to sing, the territory holder usually responds aggressively to drive the intruder out. At first, this may mean simply singing back, but can quickly escalate to full-on physical conflict.
In our experiment, we used playbacks of great tit song to mimic an intruding male. This allowed us to measure how strongly urban and rural great tits respond to intruders. We found that city birds approached the intruder mimic 35 seconds faster and almost two metres closer than rural birds. This suggests that urban great tits are both bolder and more aggressive than rural great tits.
And it’s not just great tits which are known to be more aggressive in cities. Urban song sparrows (Melospiza melodia) have also been found to be more aggressive towards each other during the breeding season.
Why urban birds should be bolder and more aggressive than their rural cousins is not yet fully understood. One possibility is that in cities, where space is limited, only the most aggressive individuals are able to hold a territory.
Animal personalities
In addition to researching levels of aggression, we also looked at how consistent males were in their aggressive displays. This consistency in the way animals behave over time is often referred to as “animal personality”.
We found rural great tits were very consistent in the way they behaved towards an intruder over two consecutive days, but urban birds showed lower levels of consistent behaviour. This could be due to the fact that urban males experience greater fluctuation in their environment on a day to day basis.
Urban bird territories are also packed closer together in cities, which means that the birds are likely to cross each others’ paths more often, increasing the chances that squabbles will break out.
The increased likelihood of aggressive encounters may also explain the lack of consistency of urban male behaviour. Animals remember fights they have had and their experience alters the way they behave the next time they encounter an opponent. Urban males are more likely to have had an aggressive encounter with males in their territory, which could then alter the way they behave towards our mimic of an intruder the following day.
As well as aggressive behaviour, being bold may also help birds survive in the city. Bolder birds have been shown to be more willing to explore new environments and to find new types of food to eat. As cities are vastly different to natural habitats, and contain many new types of food, boldness is likely to be a very useful trait for urban birds to have.
The big question which remains for ecologists is whether birds in cities are evolving to suit their new urban environment. A recent study has shown genetic differences between rural and urban great tit populations which suggests this may be the case.
Animals in urban environments face many unique challenges including finding and adapting to new sources of food, coping with almost continual human disturbance, and learning to avoid introduced predators (domestic cats I’m looking at you).
Another factor that all animals in urban habitats must learn to cope with is noise. Noise in cities comes primarily from cars and other road vehicles, but is also produced by factories, building sites, roadworks and numerous other human activities. In my own experiments I have measured noise levels in excess of 70 dB next to major city roads. For comparison, that’s as loud as a vacuum cleaner from 3 metres (10 feet) away! Even in quieter areas and at the quietest times of day urban noise rarely dips below 50 dB which is around the same level as in a bustling office.
Urban habitats are noisy places
For animals in urban environments there is almost no escape from noise. This presents a major problem for species which use sound to communicate as their calls and signals can easily be drowned out and lost in the din of the city. However, as is often the case in nature animals have found a solution to this problem and it is surprisingly simple.
In noisy environments many species, including all mammals and birds so far tested, unconsciously increase the amplitude of their vocal signals to ensure they are heard. This response to noise is known as the Lombard effect, named after its discoverer the French scientist and doctor Étienne Lombard (1869 – 1920).
Etienne Lombard (1869 – 1920). The discoverer of the Lombard effect
Surprisingly however, despite numerousstudiesoftheLombardeffect in many different species very little is known about how quickly the Lombard effect acts after sudden increases in noise. For city dwellers the ability to respond to changes in noise levels quickly may be vital as things such as passing vehicles or sporadic building or road works can cause noise levels can fluctuate wildly over the course of a day. Even in wilderness habitats noise levels can change quickly due to wind, rain or the even the calls of other animals.
The Lombard effect has been shown in every species of bird and mammal so far tested and across evolutionarily diverse groups. Figure adapted from Brumm and Zollinger (2011)
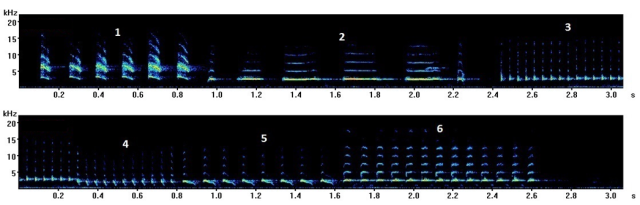
Last year my colleagues and I set out to fill this gap in our knowledge by testing how fast a songbird could exhibit the Lombard effect when noise levels suddenly changed. As a test species we used wild-type canaries. This species was perfect for our study as canaries sing a lot with little encouragement and produce really long and complex songs. These songs are composed of long strings of repeated bursts of sound known as song elements which can be grouped into different element types.
One of the canaries used in our experiment
Here you can listen to the song of one of the canaries from our experiment while the image shows a visualisation of song and song elements.
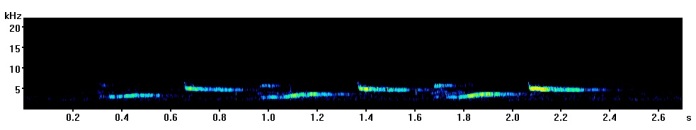
Spectrogram showing a visualisation of canary song from one bird. Each number indicates a different element type.
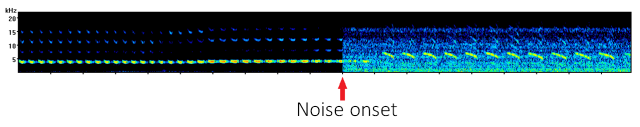
To test how quickly canaries could respond to noise we created software which would detect when a canary began to sing, then play a 20 second burst of 75 dB white noise after a random delay of between 5-10 seconds. In this way the first half a canary’s song was sung during quiet conditions while the other half was overlapped by noise which caused the bird to respond by singing louder.
In this recording you can hear how song sounds when overlapped halfway through by white noise.
An example of canary song overlapped halfway through with white noise. The noise induced the Lombard effect, forcing the canaries to sing louder
Using recordings we were able to work out the strength of the Lombard effect in our canaries by comparing how loud they sang before and after noise began.
We found that in comparison to song elements sung during quiet conditions, song elements sung during the 20 second white noise playbacks were 5.3 dB louder on average. That may not sound like much, but is actually an impressive 84 % increase in song amplitude.
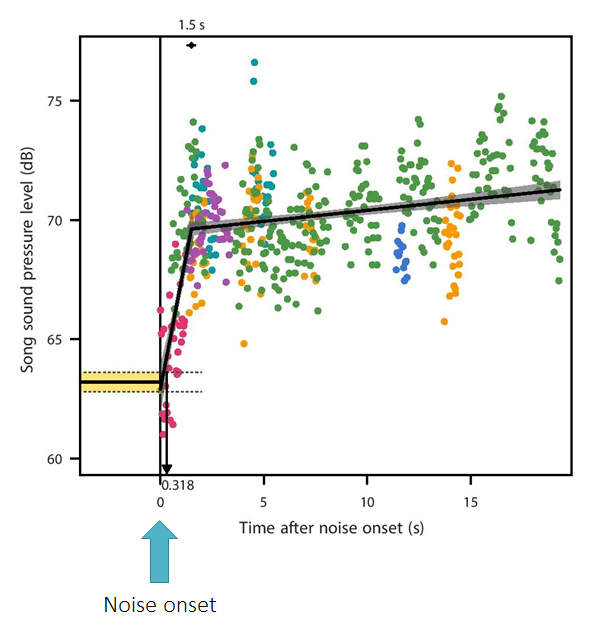
To work out how fast this increase in song amplitude occured we used a statistical technique known as broken-line regression. Using this method we were able to work out exactly how quickly the amplitude of song increased after noise began, and it was extremely fast. Our canaries were able to respond to noise extremely quickly. After just 0.32 seconds of noise exposure their song showed a statistically significant increase in amplitude.
Figure showing the speed of the onset of the Lombard effect. Before the onset of noise the mean sound pressure level (amplitude) of the song is shown by the black line with dashed lines showing the +/- 95 % confidence intervals. After noise begins the amplitude of the song rapidly increases and is significantly louder than before the onset of noise after 0.318 seconds. Each point of this figure shows the amplitude of a single song element. The different colours show elements from different recordings.
Our study shows that canaries are able to rapidly increase the amplitude of their song in response to sudden increases in background noise levels. This means that, despite fluctuating and unpredictable changes in noise levels, canaries can ensure that their songs ares till heard. This ability is likely to be particularly useful in urban areas where noise from anthropogenic sources is loud, fluctuating and unpredictable. Given that the Lombard effect is known in all other birds so far tested it is highly likely that other species also possess this ability.
This study is published in the Journal of Experimental Biology and is available here.
If you don’t have access to the journal but would like a copy of the article let me know and I will send you a copy.
As I was coming home from work yesterday there was an unmistakable feeling of spring in the air. After what feels like a very long and harsh winter (it dropped to an incredible -16°c for a few days) the sunshine and warmth is certainly welcome.
It’s not just me that’s enjoying the sunshine either. At this time of year the breeding season for great tits (Parus major) really gets going and when the sun shines the males sing as they attempt to attract females to their territories.
Great tits sing from around February to early June
For some people the first snowdrops mark the start of spring, for others the melting snow or the lengthening days. For me, great tit song signals that spring is on its way and is always a welcome sound.
Great tit song is one of the most distinctive sounds in nature. From February until early June these little birds can be heard singing in fields, parks, woodlands and even in the middle of cities from Ireland all the way to China.
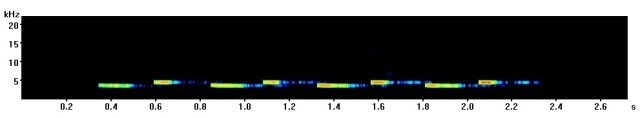
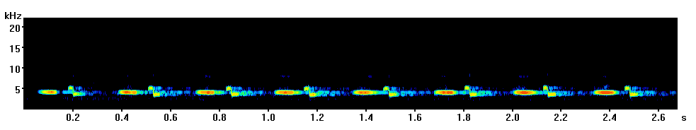
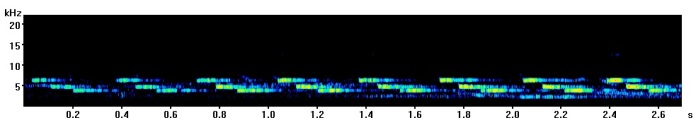
The most common great tit song contains two repeating notes, one high and one low which are said to sound a bit like “tea-cher, tea-tea-cher”. Here is an example I recorded in Derby (UK) a few years ago with a spectrogram below so you can see how it looks.
Remarkably however, this is just one of over 70 different calls and songs great tits are known to produce with individual birds having repertoires of up to eight song types. In fact, the great tit’s musical repertoire is so vast that if you hear a bird song you don’t recognise there is a good chance it’s a great tit!
Here a few different variants on the “classic” great tit call I have recorded in the past with spectrograms so you can see the difference.
Recorded in Starnberg (Germany)
Recorded in Bath (UK)
Recorded in Durham (UK)
Recorded in Leamington Spa (UK)
In the past I travelled all over the UK recording these birds as part of my research. I have recorded so many great tit songs that I have an almost Pavlovian response whenever I hear one as I feel the need to grab my microphone and start recording. This year however, all I need to do when I hear their song is look forward to longer days and better weather.
I think all who attended the meeting agreed it was a huge success. It was fantastic to spend a few days talking with other behavioural ecologists about their work and also to be able to discuss my own work with other like-minded people. Whenever I attend meetings like this I always come away with a renewed enthusiasm for what I do and with new ideas buzzing around my head for potential future research projects.
This time I did not give a talk myself, but I did present a poster showing the results of my recent work on the Lombard effect in canaries. I will write a full blog post about this soon but for those who might be interested in the poster I thought I would post it here. If you have any questions please don’t hesitate to ask!
My poster from Etho 2017. A meeting of the Ethologische Gesellschaft in Bonn.
As I mentioned in my last post I successfully defended my PhD viva (thesis defence) just over three weeks ago. As I was preparing to defend my thesis I found it helpful and often reassuring to readthe experiencesof other PhD students to get some idea of what I could expect. Of course, every viva will be different but below are some of my thoughts post-viva which I hope my be useful to others.
Preparation
In the weeks leading up to my viva I tried to prepare as best I could by re-reading my thesis and refreshing my memory of the major papers which I had cited often or based my own work upon. Apart from this I didn’t feel like I could really do much to prepare as I had no idea what questions I would be asked. At the time I found this quite stressful because I felt like I should have been working harder and doing more to prepare. Now with the benefit of hindsight I can see that I did the right things.
What my reviewers most wanted to know was that I understood and could explain my own work. Since I had spent the last 3+ years working on it this was not a problem! What I now know is that the preparation for my viva began on the very first day of my PhD. While it was helpful (and definitely recommended) to refresh my memory of the key points of my work in the weeks leading up to my viva, the real hard work had already been done over the past years.
The warm-up
On the day of my viva I got up early and read a few notes one last time over breakfast. I then headed into my university for a pre-viva chat and coffee with my supervisor. This really helped calm any nerves I had as he seemed quite confident that I would have no problems.
In the hour before my viva began I had to go and register with the secretary before being shown to the room where I would meet my reviewers. Here I had to wait outside for quite a few minutes and it was at this point that I first started to get really nervous. I think because the time waiting gave me the time to think of all the possible worst case scenarios!
Eventually, I was shown into the room where my reviewers introduced themselves and explained what the viva was and what would happen over the next few hours. I was offered drinks which had been prepared for us (tea/coffee/water) before the viva really began. These first few minutes felt like a warm-up before the real questioning began.
Full-speed
The first thing I was asked to do was to explain in broad terms what my thesis was about, why I did it and what I had discovered. This was possibly the easiest question of the whole day as I had spent so long working on my PhD that I could explain it in my sleep. The only difficulty I had was condensing more than three years of results into fifteen minutes of explanation. This meant leaving out everything but the biggest and most important elements of my work.
On a couple of occasions I found myself spending too much time explaining minor details and had to force myself to move on to more important aspects of my work. Nevertheless, this question really helped me to get into the flow of explaining my research and after a few minutes I really felt like I was firing on all cylinders.
After this warm-up question things got a lot more detailed. My reviewers went through each chapter one by one and stopped wherever they had a question about something I had written or wanted some more detail about a particular point I had raised. In almost all cases these questions were not too difficult, and in most cases they were relatively easy to answer.
However, I don’t want to suggest that there were no difficult questions. On a couple of occasions a question was put to me that I couldn’t answer very well or a criticism of my work was made which I could only agree with. During the viva this made me quite uncomfortable, but now, with the benefit of hindsight, I can see that my reviewers pushed me to look at my work from a different perspective and consider new possibilities. This is probably one of the best things to come out of my viva and will really help me as I write-up some of my chapters for publication.
Towards the end
My viva was just under three hours long. It sounds like a really long time but it really didn’t feel like it. After the first few questions it felt more like a friendly discussion that an exam and I relaxed a little bit and started to feel slightly more confident. This, along with the stress and adrenaline really helped the time to fly by.
Once both my reviewers were satisfied that they had no more questions to ask me I was asked to leave the room for a few minutes while they discussed how I did and what the outcome should be. This was probably the most nerve-wracking moment for me as I really had no idea how well I’d done or what outcome to expect. After maybe five minutes I was asked back into the room where my reviewers both seemed happy and congratulated me on passing with minor corrections. This means I have up to six months to make a few changes to my thesis which in reality should take no more than a week or two.
And relax!
The feeling of relief on being given the news that I had passed was overwhelming and I didn’t really know how to react. The corrections I need to make are only very small and my reviewers really seemed to like my work.
Once the viva was over I had a celebratory pint in the local pub with my supervisor, a reviewer and some friends. At this point I was still dazed, relieved, tired and hadn’t really full absorbed had what just happened. I think it took a full 24 hours before it really sank in that all of my work over the past years had been worth it. Often it was fun, at times it was hard, stressful and I felt like I couldn’t do it. But now it has paid off and I am happy!
If you are a PhD student about to defend your thesis and have any questions about the process please feel free to email me or leave a comment. And most of all, good luck!
The very first post on this blog was written while I was still an undergraduate at the University of Derby. Today I am happy to say that I have just successfully defended my PhD thesis on the effects of urbanisation on the behaviour of song birds. This means not only can I order a new bank card with “Dr.” on it, but also that my time as a student has finally come to an end.
The first draft of my PhD thesis just before I submitted it.
The question I have to ask myself now is what next?
Right now I am working at the Max Planck Institute for Ornithology in Germany and am loving it. I work with a really fantastic research group and I hope I will be able to stay here a while yet to pursue some of my research ideas. After that, I have no idea! I am considering several possibilities including continuing in academia long-term, working for conservation/environmental charities or applying for jobs in government environmental agencies. If you have completed a PhD I would love to hear in the comments what you did next.
As for this blog, I hope to revive it after its long and unplanned hiatus. As all PhD students know, the final year is the most intense and personally challenging. In the last year I have moved to Germany, written my thesis and published a paper. amongst all this I didn’t find the time for my poor blog! Now, however, I have a little more time and I plan to use it to write a series of blog posts on my PhD experience. Both the good and the bad. If there is anything you would like to know about the PhD experience please let me know below!
I also plan to write about whatever I do next. Although it is still quite unclear at the moment I am optimistic for the future and looking forward to the next challenge. The end of being a student me is really just the beginning of the next stage in my life and I can’t wait!
In 1800 only 3% of the world’s population lived in urban areas, yet as the industrial revolution picked up pace in the early 18th and 19th centuries the number of people moving from the countryside to work in the newly industrialised cities soared. By 1950 29% of the world’s population were living in cities and by 1985 this had grown to 42% while in 2025 it is estimated to that it will be over 60%1. That is just 10 years from now. In highly developed nations such as those in Western Europe and North America the 50% threshold has already been surpassed which means that if you live in a western nation and do not live in a city you are in a minority.
Across the globe cities are expanding.
It’s not just where people choose to live that is rapidly changing either, but how many of us there are. The human population is currently expanding more rapidly than ever before and has grown from just a few hundred million people less than a thousand years ago to over 7 billion today, and this number is still rising2. As the human population grows cities are expanding quickly to meet the need for additional housing while the surrounding countryside is farmed and developed ever more intensely to provide us with the food, water and other resources we need to live.
Human population growth from around 5 million people in 8000 BCE to over 7 billion today. (figure from Keinen and Clark 2012).
Naturally, many people are concerned about how we are going to continue to feed and clothe ourselves as populations continue to expand, but there is also a growing concern among many about how such huge numbers of people are affecting the environment and the animals that inhabit it.
For animals which depend on natural habitats such as forests, meadows, or wetlands to survive, the growth of urban areas often spells bad news as these habitats are removed and paved over to make way for new suburbs, factories and roads. In the UK numerous species have declined for just this reason. For example, the bittern, a close relative of herons, was once widespread in reedbeds and wetlands across the UK but is now confined to a tiny area of the south-east after its habitats were drained to make way for agriculture and urban developments.
While many species cannot survive in urbanised areas, others are able to tolerate moderate levels of urbanisation and may continue living within cities despite drastic changes to their habitats. Life in the city is not without its challenges however, even for the most adaptable and resilient of species. Cities typically contain different threats to rural areas such as an abundance of cats which are responsible for killing huge numbers of birds and small mammals4, and high levels of chemical5, light6 and noise pollution7 which all have negative impacts.
This all sounds bad, and it really is, but while many species suffer badly from the effects of increasing urbanisation and habitat loss, there are a few species that have been able to adjust remarkably well to life in urban areas. One group that has been particularly well-studied in this regard are the songbirds and over the past 15 years or so, biologists have discovered some fascinating behavioural adaptations which have allowed some species to become successful city dwellers.
Great tits are one species which are adapting to city life.
One of the most notable features of cities across the world is that they are incredibly noisy places. With heavy traffic, building sites, aircraft flying overhead and all manner of other sounds and distractions it’s a miracle anyone can hear anything at all. For songbirds however, all this noise is more than just a distraction, it can seriously affect their chances of finding mates and successfully reproducing and for males it is likely to affect how well they can defend their territories against rivals.
While we may find bird song pleasant to listen to (or annoying depending on how early in the morning it is), for songbirds it has a serious purpose. Males sing during the breeding season to attract females8 and to signal to other males that their territory is occupied and should not be entered9.
City noise can overlap and interfere with these signals making communication among birds difficult and unreliable. The background noise of a city is typically continuous low rumble concentrated at around 2kHz in frequency. Unfortunately for many birds this overlaps neatly with the frequency of their songs and this can make it difficult for other birds to hear them as they do not stand out from the irrelevant background noise.
Clearly this is a problem for birds which rely on song to communicate, yet research has revealed that birds have ways of overcoming this problem and one of them is to increase the frequency at which they sing so that their songs literally rise above the background noise and can be clearly heard.
Sonograms of rural (a) and urban (c) great tit song compared to the background noise of a city (b). You can clearly see that the urban song is a higher frequency than rural song and above the frequency of the city noise. (from Mockford and Marshall, 2009).
Evidence that birds sing at higher frequencies in noisy cities than they do in quieter rural sites has now been found in numerous species including great tits (Parus major)7, blackbirds (Turdus merula)10, European robins (Erithacus rubecula)11 and song sparrows (Melospiza melodia)12.
In great tits the difference in song frequency between urban and rural sites has been measured at 478Hz13 and tests have shown that this is enough to substantially improve the distance over which song can travel in urban environments before it degrades and becomes inaudible14.
Birds may also face challenging noisy conditions in natural environments too such as where running water or wind creates high levels of low-frequency noise and these naturally noisy sites have allowed scientists to confirm that it really is the noise in cities and not some other factor which is causing city birds to sing at high frequencies. Biologists Henrik Brumm and Hans Slabbekoorn recorded the songs of white-throated dippers (Cinclus cinclus) living around noisy fast flowing streams in Scotland and found that they call at frequencies well above that of the background noise and higher than usual for this species suggesting that dippers in this area have adapted their calls to suit their noisy habitat15.
Dippers close to noisy streams sing at a higher frequency than the background noise so their songs can be heard (from Brumm and Slabbekoorn, 2005).
The effect of natural background noise on song frequency has also been shown in African little greenbuls (Andropadus virens) which sing at a higher frequency in areas where the rainforest is merging with open grasslands (known as ecotone forests) than they do deep within the rainforest itself16. Analysis of these two habitats revealed that the background noise in the rainforest is largely concentrated at higher frequencies while in ecotone forests there is more low-frequency noise. By singing at a lower frequency little greenbuls within the rainforest can ensure that their song does not overlap with the higher frequency background noise found in rainforests, while by singing at a lower frequency little greenbuls in ecotone forests avoid the lower frequency background noise in their habitat.
The evidence that birds change the frequency of their songs as an adaptation to noisy conditions may seem quite conclusive but not everyone agrees. An alternative explanation for the observed frequency shifts is that higher frequency song is actually just an unavoidable and possibly unimportant side-effect of singing more loudly, and it is higher volume, not frequency, which allows birds in noisy environments to overcome the background noise17.
In support of this argument Erwin Nemeth and Henrik Brumm of the Max Planck Institute for Ornithology in Germany found that the typical increases in song frequency found in great tits and blackbirds may be too low to substantially improve signal transmission whereas small increases in song amplitude were found to increase the distance over which a bird’s song could be detected much more effectively18.
But why should song frequency increase when birds sing more loudly? Nemeth and Brumm suggest two possibilities. Firstly, the increase in frequency observed in songs in noisy environments could be a side-effect of what is known as the Lombard effect (named after the French scientist Étienne Lombard) in which animals unconsciously increase the volume and frequency of their calls when the level of the background noise rises.
Nightingales in Berlin sing loudly to ensure that they are heard.
The Lombard effect is known to occur in humans (this is why it might feel like you have to shout to be heard at loud parties) and has also been shown in both lab and field studies of songbirds. Lab experiments on elegant crested tinamous (Eudromia elegans)19, and budgerigars (Melopsittacus undulates)20, have shown that these species both sing more loudly and at a higher frequency when background noise increases and the same result has been shown in the field in a study of Nightingales (Luscinia megarhynchos) in Berlin21.
A second reason why song frequency may increase when birds sing more loudly is that both the volume and frequency of bird songs depend on the same song producing organ which could limit how well birds can independently control frequency and volume. In birds this organ is the syrinx which is the bird equivalent of the mammalian larynx or voice box and is located at the base of the windpipe connected to the lungs. Birds produce song by forcing air at high pressure from the lungs through the syrinx causing membranes to vibrate creating sound. This sound can then be modified using numerous tiny muscles which alter the shape and tension of the sound producing membranes.
However, past studies of the avian vocal system have shown that without these tiny muscles altering the structure of the sound, both the frequency and amplitude of bird song unavoidably increase together. In other words, when birds sing louder they cannot help but also sing at a higher frequency22.
Of course, this may be totally irrelevant if birds are able to use muscles to independently control the frequency and volume of their songs but there is evidence to suggest that the frequency and volume of bird songs really are closely intertwined. One of the clearest examples of this comes from a study by Nemeth and his colleagues at the Max Planck institute who recorded blackbirds singing in sound-proof chambers and showed that volume and frequency really were strongly correlated17. When blackbirds sing more loudly they also sing at a higher frequency and this may be totally involuntary. Similar results have been found in other species including zebra finches (Taeniopygia guttata)23 and song doves (Streptopelia risoria)24 suggesting that this pattern may be widespread in birds as a whole.
When blackbirds increase the volume of the song they also sing at a higher frequency. (image credit Juan Emilio).
It has become very clear over the past few years that urban noise is causing bird song to change however, opinion is still divided on whether it is the frequency or amplitude changes that are most important to improving song transmission in noisy environments. It is possible that both have important roles to play in helping birds to adapt to noisy urban areas and hopefully future research will provide an answer to this question.
The study of how urban noise affects bird song is a very active area of research and there are many unresolved questions which are likely to be answered in the next few years. Most importantly we need to find out what the long-term impacts of urban noise are on bird populations. Although many species of birds do seem to be able to adapt to noise we do not know how the dramatic changes we are causing to their environments will affect them in the long-term. Furthermore, many species are not able to adapt to urban areas for numerous possible reasons. They may not possess the behavioural flexibility to cope with new environments or not they might not be physiologically capable of changes their songs or behaviour. That is why studies those discussed here matter, we are changing the planet in ways which have never been seen before and we know that many species are suffering as a result. The first step to protecting animals from these changes is to understand how they are affected and that is just what these studies aim to do.
References
1. Kegel, B (2014).Tiere in der Stadt: Eine Naturgeschichte. Köln: DuMont Buchverlag. (In German).
2. Keinan, A., & Clark, A. (2012). Recent Explosive Human Population Growth Has Resulted in an Excess of Rare Genetic Variants. Science, 336 (6082), 740-743 DOI: 10.1126/science.1217283
3. Barnosky AD, Matzke N, Tomiya S, Wogan GO, Swartz B, Quental TB, Marshall C, McGuire JL, Lindsey EL, Maguire KC, Mersey B, & Ferrer EA (2011). Has the Earth’s sixth mass extinction already arrived?. Nature, 471 (7336), 51-7 PMID: 21368823
4. van Heezik, Y., Smyth, A., Adams, A., & Gordon, J. (2010). Do domestic cats impose an unsustainable harvest on urban bird populations?. Biological Conservation, 143 (1), 121-130 DOI: 10.1016/j.biocon.2009.09.013
5. Liker A, Papp Z, Bókony V, & Lendvai AZ (2008). Lean birds in the city: body size and condition of house sparrows along the urbanization gradient. The Journal of animal ecology, 77 (4), 789-95 PMID: 18479344
7. Slabbekoorn, H., & Peet, M. (2003). Ecology: Birds sing at a higher pitch in urban noise. Nature, 424 (6946), 267-267 DOI: 10.1038/424267a
8. Baker, M., Bjerke, T., Lampe, H., & Espmark, Y. (1986). Sexual Response of Female Great Tits to Variation in Size of Males’ Song Repertoires. The American Naturalist, 128 (4) DOI: 10.1086/284582
9. Krebs, J., Ashcroft, R., & Webber, M. (1978). Song repertoires and territory defence in the great tit. Nature, 271 (5645), 539-542 DOI: 10.1038/271539a0
10. Nemeth, E., & Brumm, H. (2009). Blackbirds sing higher-pitched songs in cities: adaptation to habitat acoustics or side-effect of urbanization? Animal Behaviour, 78 (3), 637-641 DOI: 10.1016/j.anbehav.2009.06.016
11. McLaughlin, K., & Kunc, H. (2012). Experimentally increased noise levels change spatial and singing behaviour. Biology Letters DOI: 10.1098/rsbl.2012.0771
13. Mockford, E., & Marshall, R. (2009). Effects of urban noise on song and response behaviour in great tits. Proceedings of the Royal Society B: Biological Sciences, 276 (1669), 2979-2985 DOI: 10.1098/rspb.2009.0586
14. Mockford, E., Marshall, R., & Dabelsteen, T. (2011). Degradation of Rural and Urban Great Tit Song: Testing Transmission Efficiency. PLoS ONE, 6 (12) DOI: 10.1371/journal.pone.0028242
15. Brumm, H., & Slabbekoorn, H. (2005). Acoustic communication in noise. Advances in the Study of Behavior, 35, 151-209 DOI: 10.1016/S0065-3454(05)35004-2
16. Slabbekoorn H, & Smith TB (2002). Habitat-dependent song divergence in the little greenbul: an analysis of environmental selection pressures on acoustic signals. Evolution; international journal of organic evolution, 56 (9), 1849-58 PMID: 12389730
17. Nemeth, E., Pieretti, N., Zollinger, S., Geberzahn, N., Partecke, J., Miranda, A., & Brumm, H. (2013). Bird song and anthropogenic noise: vocal constraints may explain why birds sing higher-frequency songs in cities. Proceedings of the Royal Society B: Biological Sciences, 280 (1754), 20122798-20122798 DOI: 10.1098/rspb.2012.2798
18. Nemeth, E., & Brumm, H. (2010). Birds and Anthropogenic Noise: Are Urban Songs Adaptive?. The American Naturalist, 176 (4), 465-475 DOI: 10.1086/656275
19. Schuster, S., Zollinger, S., Lesku, J., & Brumm, H. (2012). On the evolution of noise-dependent vocal plasticity in birds. Biology Letters, 8 (6), 913-916 DOI: 10.1098/rsbl.2012.0676
20. Osmanski, M., & Dooling, R. (2009). The effect of altered auditory feedback on control of vocal production in budgerigars (Melopsittacus undulatus). The Journal of the Acoustical Society of America, 126 (2) DOI: 10.1121/1.3158928
21. Brumm, H. (2004). The impact of environmental noise on song amplitude in a territorial bird. Journal of Animal Ecology, 73 (3), 434-440 DOI: 10.1111/j.0021-8790.2004.00814.x
22. Titze, I. R. (1994). Principles of voice production (pp. 279-306). Englewood Cliffs: Prentice Hall.
23. Cynx J, Lewis R, Tavel B, & Tse H (1998). Amplitude regulation of vocalizations in noise by a songbird, Taeniopygia guttata. Animal behaviour, 56 (1), 107-13 PMID: 9710467
24. Elemans, C., Zaccarelli, R., & Herzel, H. (2008). Biomechanics and control of vocalization in a non-songbird Journal of The Royal Society Interface, 5 (24), 691-703 DOI: 10.1098/rsif.2007.1237
The last few months have been a blur of conferences, workshops. I attended a fantastic two week course on sensory ecology in Sweden (read about that here and here), I went to London and presented a poster of my work at the annual meeting of the Association for the Study of Animal Behaviour and have recently returned from the Joint meeting of the British Ecological Society and the Société Française d’Écologie in Lille. Time has been flying by and now Christmas is upon us but I wanted to write at least one more post for 2014 so here it is. These are four facts about my wonderful study species the great tit (Parus major).
My study species.
Every great tit has a unique personality
It’s true, you might think that a bird is just a bird and they are all the same but research has shown that this is not the case. In fact, many species of birds including great tits are now known to have distinct personalities and behave differently to one another, just like humans do. Researchers at Groningen University in the Netherlands found that when they exposed adult great tits to a completely new environment which they had never experienced before some birds were confident and bold and explored their new environments quite happily, while others were much more cautious and did not adapt quickly to the unfamiliar surroundings. Further tests have shown that birds which are bold or shy the first time they are tested tend to stay that way in repeated experiments. These results suggest that personality types are probably fixed characteristics which do not easily change. So, for anyone who’s ever said that animals have personalities, you were right. And it’s not just great tits either; research over the past few years has identified distinct personalities in a huge number of species including mammals, birds, insects and even anemones!
Different personalities. Great tits are not all the same. Image credit: Per Tillmann.
City great tits just aren’t as colourful as their countryside cousins
With their bright yellow breast feathers and marked and glossy black plumage great tits are hard to miss at the best of times and are particularly conspicuous during the breeding season when the males are singing their hearts out trying to attract mates. The purpose of the great tits striking colouration is not fully understood but it is likely that females are most attracted to those males which have the brightest and boldest feathers. It is unfortunate then for city birds that their feathers just aren’t as bright as their rural counterparts. A likely explanation for this is that the same carotenoid compound which is used to create the bright yellow colour in great tits feathers is also an anti-oxidant which reduces the physiological stress caused by oxidising pollutants in the environment. As urban birds are known to be exposed to much higher levels of pollutants than rural birds it may be that urban birds are forced to divert carotenoids away from their feathers and use them instead to reduce the levels of stress and cell damage that oxidising compounds can cause. Evidence of this comes from a study at Göteborg University in Sweden which found that the increased oxidative stress faced by urban birds correlated well with reduced levels of carotenoid pigments in their feathers.
Great tits can be incredibly aggressive
They might look cute and friendly but in reality great tits can be incredibly violent, and particularly so during the spring and summer when males are competing for the best breeding territories. During this period male great tits mark their territories by singing at regular intervals and this signal tells other males to move on and find an empty territory of their own. This works most of the time but not always. Sometimes males deliberately invade territories and may attempt to mate with the resident female or even try to force the current territory holder out and take over completely. If this happens the first thing the resident male will do is to sing frequently and loudly in an attempt to show their strength and to drive the intruder away. Should this not work things typically escalate quickly as the resident male flies back and forth across the intruder before coming into full-on physical confrontation. It is not common to see fights in the wild as invaders will usually leave before it comes to actual violence. It does happen sometimes though and you can see the result in these incredible videos.
Great tits sing at higher frequencies in cities than in rural areas
Cities are notoriously noisy places full of cars, people and factories, with aircraft flying overhead and noise from building sites, road repair crews or similar urban development projects. This level of noise is something which wild animals have only had to face in the last century or so, and for species which communicate using sound it could make life in cities very difficult as their vocal signals are easily lost amongst the clatter and din of urban life. Great tits are a good example of a species which might be expected to fare badly in urban habitats as the males rely on communicating by song to defend their territories, show aggression towards rivals and attract mates. To make matters worse, the typical song of a great tit lies in exactly the same frequency range as the background noise of most cities and this means that when a great tit sings its song is quickly swamped by urban noise and cannot be heard by other birds. Yet despite this, great tits are one of the most common song birds seen in city parks and at garden bird feeders so how do they manage to cope with the noise problem? The answer is simple but ingenious. Urban great tits have adjusted their typical song frequency so that they now sing around 350Hz higher. That might not sound like much but experiments have shown that this small adjustments means that the song of urban birds travels above the low-frequency rumble of urban noise and can still be heard by other birds, even at long distances. Genius!
When there are high background noise levels great tits adapt by singing at a higher pitch. Figure from Slabbekoorn and Peet (2003).
References
Every great tit has a unique personality
Dingemanse, N. (2002). Repeatability and heritability of exploratory behaviour in great tits from the wild Animal Behaviour, 64 (6), 929-938 DOI: 10.1006/anbe.2002.2006
City great tits just aren’t as colourful as their countryside cousins
Isaksson, C., Örnborg, J., Stephensen, E., & Andersson, S. (2005). Plasma Glutathione and Carotenoid Coloration as Potential Biomarkers of Environmental Stress in Great Tits EcoHealth, 2 (2), 138-146 DOI: 10.1007/s10393-005-3869-5
Great tits sing at higher frequencies in cities than in rural areas
Slabbekoorn, H., & Peet, M. (2003). Ecology: Birds sing at a higher pitch in urban noise Nature, 424 (6946), 267-267 DOI: 10.1038/424267a
It’s the start of a new month and that means it’s time again for the Carnival of Evolution and this time it’s a special one, the 75th edition! So all aboard as we journey once more through some of the best evolution based blog posts from around the web.
Our first stop is the History of Evolution so step out and enjoy the view, there are many good things to see here.
Did these two species evolve in sympatry? Mycocepurus goeldii (left) and Mycocepurus castrator (right) are sister species which occupy the same geographic area (image credit: Christian Rabeling, University of Rochester).
And that’s it for evolutionary ecology this time, now our journey must continue as we head into the wonderful world of Evolutionary Theory.
Ben Haller talks about the interactions between empirical and theoretical researchers and asks Should theoretical ideas drive new empirical work to look for the patterns and outcomes predicted by theoretical models? Or should pure “natural history” observations of the real world drive new theoretical work to explain the patterns and outcomes observed?
Our journey is almost at an end but not quite! Hold tight, we have three more posts to go.
And now we come to the end of the line. We hope you had a pleasant journey and will travel with us again soon. The next Carnival will be held in October at Eco-Evolutionary Dynamics, if you would like to submit anything you can submit it on the Facebook page. You can keep up with the Carnival of Evolution via Facebook and Twitter.
A few months ago during the coffee break at an animal behaviour conference I was talking to a colleague about her research when she told me that to suggest that animals could feel fear or be afraid was anthropomorphism, the mistake of assigning of uniquely human characteristics to other animals. This view is not at all uncommon among practicing scientists and the term anthropomorphism is often extended to include a whole range of behavioural traits and emotions such as impatience, joy, expectation, boredom, anger, happiness or sadness, and yet there is good evidence that these emotions are not unique to humans. For example, dogs have been shown to exhibit jealousy, elephants have empathy, and Capuchin monkeys get visibly angry when treated unfairly as this video shows.
The possibility than animals can think for themselves is also often questioned and yet we know that some animals such as chimps and dolphins have a sense of self. There are also examples of animal behaviours that surely require some degree intelligence and forward planning such as innovative tool use by chimps and deception by ravens.
Deep in thought. Do chimps think like we do?
One of the main arguments against using anthropomorphic language to describe animal behaviours is that there is no way to know how an animal is really feeling, we can only describe what it looks like it’s feeling but not what’s actually happening inside it’s head. But the same is also true of humans, yet no one would question the use of anthropomorphic language to describe human behaviours.
If a person says they are excited we don’t actually know that what they feel as excitement is the same as what you or I feel as excitement, to them it may be a very different thing. All we can do is observe how that person acts and behaves and decide for ourselves if that matches up with our interpretation of what excitement is. The same is true of any emotion. If I say I am or happy or bored how could you tell that what I feel as happiness or boredom is the same as what you or anyone else feels as those things? At some point we have to use our subjective judgement to decide how a person is feeling. Do their actions match our expectations for a happy person? Then we can say they are happy. Do they behave as if they are sad? Then we can say they are sad. If this applies to humans then surely it can also apply to animals, at least in some cases.
I am not suggesting that we should abandon all caution and start using anthropomorphic terms carelessly. What I am suggesting is that so long as we clearly define our terms we should be able to use words like ‘afraid’ or ‘excited’ to describe animal behaviours when those terms well match what we see. If I want to describe anger in animals I should be able to use the word anger so long as I clearly state what I mean by the term and the behaviour that I am describing well matches what most of us would recognise as anger.
Of course, there are cases where using anthropomorphic terms really isn’t appropriate. If I read a paper that described ‘angry’ aphids or ‘jealous’ earthworms I would be dubious that those animals really could feel those emotions. In other cases I would have much less of a problem. Can chimps get angry? I think so. Are rats afraid of predators? It seems likely.
For many scientists, including people who I work with, the fear of using anthropomorphic language seems deeply ingrained and I think this affects how we view the behaviour of animals. Not all animals are mindless automatons that blindly follow their pre-programmed instincts. Many animals, especially among the vertebrates, have complex behaviours and emotions which are best described using the same terms we use for those things in human animals. I think it is time we started describing animals behaviours exactly as we see them. We must define our terms and we must be clear but so long as we are there should be little problem to this approach.
For those that still doubt that animals have can have thoughts and emotions like ours I recommend this TED talk.









 Remarkably however, this is just one of
Remarkably however, this is just one of 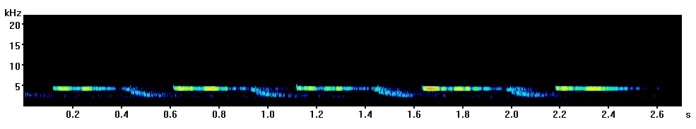







![Nachtigall_%28Luscinia_megarhynchos%29-2[1]](https://ecologicablog.files.wordpress.com/2015/02/nachtigall_28luscinia_megarhynchos29-21.jpg)
![1280px-Turdus_merula_-Gran_Canaria%2C_Canary_Islands%2C_Spain-8_%282%29[1]](https://ecologicablog.files.wordpress.com/2015/02/1280px-turdus_merula_-gran_canaria2c_canary_islands2c_spain-8_282291.jpg)









